
Maize (Zea mays L.) is the world’s leading crop and is widely cultivated as cereal grain that was domesticated in Central America. It is one of the most versatile emerging crops having wider adaptability. Globally, maize is known as queen of cereals because of its highest genetic yield potential. Maize is the only food cereal crop that can be grown in diverse seasons, ecologies and uses. Beside this maize have many types like normal yellow/ white grain, sweet corn, baby corn, popcorn, waxy corn, high amylase corn, high oil corn, quality protein maize, etc. Apart from this, maize is an important industrial raw material and provides large opportunity for value addition.
- Taxonomy, Origin and Evolution
1. A. Taxonomy
Maize belongs to the tribe Maydeae of the grass family Poaceae. “Zea” was derived from an old Greek name for a food grass. The genus Zea consists of four species of which Zea mays L. is economically important. The other Zea sp., referred to as teosinte, is largely wild grass native to Mexico and Central America. The number of chromosomes in Zea mays is 2 n = 20. Tribe Maydeae comprises seven genera which are recognized, namely Old and New World groups. Old World comprises Coix (2n = 10/20), Chionachne (2n = 20), Sclerachne (2n = 20), Trilobachne (2n = 20) and Polytoca (2n = 20), and New World group has Zea and Tripsacum. It is generally agreed that maize phylogeny was largely determined by the American genera Zea and Tripsacum, however it is accepted that the genus Coix contributed to the phylogenetic development of the species Zea mays.
| Kingdom | Plantae |
| Division | Magnoliophyta |
| Class | Liliopsida |
| Order | Poales |
| Family | Poaceae |
| Genus | Zea |
| Species | mays |
- B. Origin and Evolution
How corn evolved and what are its progenitor species is still a matter of investigation. Archaeological and molecular data indicates that modern maize was domesticated from annual teosinte (Zea mays ssp. parviglumis) in southern Mexico between 6,600 and 9,000 years ago. Selection soon followed: favourable alleles at loci controlling plant morphology and kernel nutritional quality were fixed at least 4,400 years ago, and further selection by native Americans facilitated maize adaptations to varied environments. - Developmental Stages (Life Cycle)
Typical corn plants develop 18 to 22 total leaves, silk appears about 55 days after emergence, and mature in around 125 days after emergence (Ritchie et al., 1993). The specific time interval, however, can vary among hybrids, environments, planting date, and location. The length of time between each growth stage, therefore, is dependent upon these circumstances. For example, an early maturing hybrid may produce fewer leaves or progress through the different growth stages at a faster rate than described here. In contrast, a late- maturity hybrid may develop more leaves and progress through each growth stage at a slower pace.
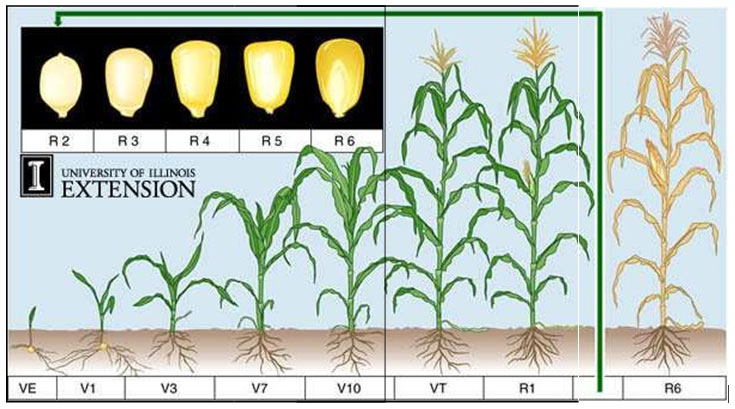
Fig. 2. Different developmental stages in plants. (R1-R6= Different growth stages in seed),
(Ritchie et al., 1993)
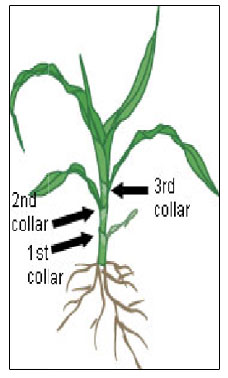
The vegetative stages of maize are identified by the number of collars present on the corn plant. The leaf collar is the light-colored collar-like “band” located at the base of an exposed leaf blade, near the spot where the leaf blade comes in contact with the stem of the plant. Leaves within the whorl, not fully expanded and with no visible leaf collar are not included. For example, a plant with 3 collars would be called a V3 plant, however, there may be 6 leaves showing on the plant.
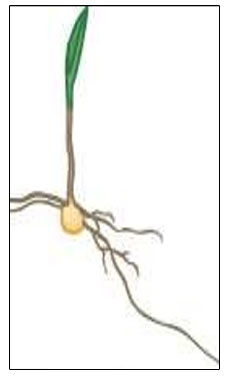
VE – Emergence
Coleoptile reaches the soil surface and exposure to sunlight causes elongation of the coleoptile and mesocotyl to stop. The growing point, located just above the mesocotyl, is about 0.75 inches below the soil surface. Embryonic leaves rapidly develop and grow through the coleoptile tip. Seminal root growth begins to slow and nodal roots are initiated at the crown.
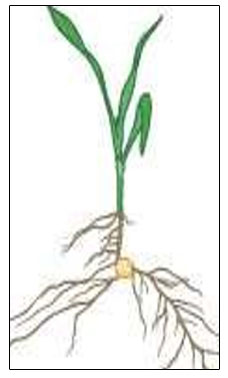
V1 – First leaf collar
Lowermost leaf (short with rounded tip) has a visible leaf collar. Nodal roots begin elongation.
V3 – Third leaf collar
The growing point remains below the soil surface as little stalk elongation has occurred. Lateral roots begin to grow from the nodal roots and growth of the seminal root system has ceased. All leaves and ear shoots that the plant will produce are initiated at this stage. Since the growing point remains below the soil surface, cold soil temperatures may increase the time between leaf stages, increase the total number of leaves formed, delay tassel formation, and reduce nutrient uptake.
V7 – Seven leaf collar
During the V7 and V8 growth stages the rapid growth phase and kernel row determination begins. Senescence of lower leaves may occur if plant is stressed, but must still be counted when staging plants.
V10 – Ten leaf collar
At the V9 and V10 growth stages the stalk is in a rapid growth phase accumulating dry matter as well as nutrients. The tassel has begun growing rapidly as the stalk continues to elongate. Many ear shoots are easily visible when the stalk is dissected.

VT – Tasseling
Initiation of the VT stage begins when the last branch of the tassel is visible and silks have not emerged. This stage begins about 2-3 days before silk emergence. The plant is almost at its full height and pollen shed (anthesis) begins. Pollen shed typically occurs in the morning or evening.

R1 – Silking
This stage begins when any silk is visible outside the husk. Falling pollen grains are captured by the silk and grow down the silk over a 24-hour period ultimately fertilizing the ovule. The ovule becomes a kernel. It takes upwards of three days for all silks on a single ear to be exposed and pollinated. The number of fertilized ovules is determined at this stage. If an ovule is not fertilized, it will not produce a kernel and it eventually degenerates.

R6 – Physiological Maturity
Occurring approximately 45-50 days after silking, all kernels on the ear have attained maximum dry weight. A black or brown layer has formed where the kernel attaches to the cob, indicating physiological maturity has been attained. The stalk of the plant may remain green, but leaf and husk tissue has lost its green color at this stage. Kernel moisture content ranges from 30- 35% at this stage, with much variation among hybrids and environmental conditions.
3. Botanical features
Maize is a tall, determinate, annual C4 plant varying in height from <1 to >4 meters producing large, narrow, opposing leaves, borne alternately along the length of a solid stem. The botanical features of various plant parts are as follows:
Root: Normally maize plants have three types of roots, (i) seminal roots – which develop from radical and persist for long period; (ii) adventitious roots, fibrous roots developing from the lower nodes of stem below ground level which are the effective and active roots of plant; and (iii) brace or prop roots, produced by lower two nodes. The roots grow very rapidly and almost equally outwards and downwards. Suitable soils may allow corn root growth up to 60 cm laterally and in depth.
Stem: The stem generally attains a thickness of three to four centimeters. The internodes are short and fairly thick at the base of the plant; become longer and thicker higher up the stem, and then taper again. The ear bearing internode is longitudinally grooved, to allow proper positioning of the ear head (cob). The upper leaves in corn are more responsible for light interception and are major contributors of photosynthate to grain.
Flower: The apex of the stem ends in the tassel, an inflorescence of male flowers and the female inflorescences (cobs or ears) are borne at the apex of condensed, lateral branches known as shanks protruding from leaf axils. The male (staminate) inflorescence, a loose panicle, produces pairs of free spikelets each enclosed by a fertile and a sterile floret. The female (pistillate) inflorescence, a spike, produces pairs of spikelets on the surface of a highly condensed rachis (central axis, or “cob”). The female flower is tightly covered over by several layers of leaves, and so closed in by them to the stem that they don’t show themselves easily until emergence of the pale-yellow silks from the leaf whorl at the end of the ear. The silks are the elongated stigmas that look like tufts of hair initially and later turn green or purple in color. Each of the female spikelets encloses two fertile florets, one of whose ovaries will mature into a maize kernel once sexually fertilized by wind-blown pollen.
Floral Biology
Maize is a monoecious plant, that is, the sexes are partitioned into separate pistillate (ear), the female flower and staminate (tassel), the male flower. The main shoot terminates in a staminate tassel. Maize is generally protandrous, that is, the male flower matures earlier than the female flower. Within each male flower spikelet, there are usually two functional florets, although development of the lower floret may be delayed slightly in comparison to the upper floret. Each floret contains a pair of thin scales i.e. lemma and palea, three anthers, two lodicules and rudimentary pistil. Pollen grains per anther have been reported to range from 2000 to 7500. Within an average of 7000 anthers per tassel and around 3500 pollen grains per anther, each tassel could produce around 2.45 crores of pollen grains. In terms of the ratio of pollen grains produced per ovules fertilized, it appears that since each ear requires about 1000 pollen grains for fertilization, there are about 20,000 pollen grains per kernel in excess of what is actually needed if pollination were 100 percent efficient. The pollen grains are very small, barely visible to the naked eye, light in weight, and easily carried by wind. The wind-borne nature of the pollen and protandry lead to cross- pollination.
The female flower initially small but protuberances soon into form of rows. The basal protuberances are formed first and development advances towards the tip of the ears. The part above the attachment of the carpel develops a single sessile ovule, which consists of a nucellus with two integuments or rudimentary seed coats. The united carpel’s, which will form the ovary wall or pericarp of the mature kernel, grow upward until they completely enclose the ovule. The two anterior carpel, which face the ear tip, form outgrowths, which develop into the style i.e. into long thread, known as silks. Silks are covered with numerous hairs, trichomes which form an angle with the silk where pollen grains are harbored. The base of the silk is unique, as it elongates continuously until fertilization occurs. The cobs bear many rows of ovules that are always even in number. The female inflorescence or ear develops from one or more lateral branches (shanks) usually borne about half-way up the main stalk from auxiliary shoot buds. As the internodes of the shanks are condensed, the ear remains permanently enclosed in a mantle of many husk leaves. Thus, the plant is unable to disperse its seeds in the manner of a wild plant and instead it depends upon human intervention for seed shelling and propagation.
Grain: The individual maize grain is botanically a caryopsis, a dry fruit containing a single seed fused to the inner tissues of the fruit case. The seed contains two sister structures, a germ which includes the plumule and radical from which a new plant will develop, and an endosperm which will provide nutrients for that germinating seedling until the seedling establishes sufficient leaf area to become autotrophy. The germ is the source of maize “vegetable oil” (total oil content of maize grain is 4% by weight). The endosperm occupies about two thirds of a maize kernel’s volume and accounts for approximately 86% of its dry weight. The endosperm of maize kernels can be yellow or white. The primary component of endosperm is starch, together with 10% bound protein (gluten).
Mating Systems
Pollination occurs when these new moist silks catch the falling pollen grains. In maize, the pollen shed is not a continuous process and usually begins two to three days prior to silk emergence and continues for five to eight days. The silks are covered with fine, sticky hairs which serve to catch and anchor the pollen grains. Pollen shed stops when the tassel is too wet or too dry and begins again when temperature conditions are favorable. Under favorable conditions, pollen grain remains viable for only 10 to 18 hours. Cool temperatures and high humidity favor pollen longevity. Under optimal conditions the interval between anthesis and silking is one to two days. About 95% of the ovules are cross-pollinated and about 5% are self-pollinated (Sleper & Poehlman 2006), although plants are completely self-compatible.
4.Type of corn
Corn variation may be artificially defined according to kernel type likes: dent, flint, flour, sweet, pop and pod corn etc. Except for pod corn, these divisions are based on the quality, quantity and pattern of endosperm composition in the kernel and are not indicative of natural relationships (Brown and Darrah, 1985).
Dent Corn
Dent corn is characterized by the presence of corneous, horny endosperm at the sides and back of the kernels, while the central core is a soft, floury endosperm extending to the crown of the endosperm. It collapses to produce a distinct indentation on drying. Degree of denting varies with its genetic background. Dent corn is used primarily as animal food, but also serves as a raw material for industry and as staple food. It is still an important human food and industrial material, entering into many specialized products via the dry- or wet-milling industry. However, white dent often receives a premium price in the dry milling industry, where it utilized for certain human food products because of its whiter starch.
Flint Corn
The flint corns mostly have a thick, hard, vitreous (glassy) or corneous endosperm layer surrounded by small, soft granular center. The relative amounts of soft and corneous starch, however, vary in different varieties. Generally, the kernels are smooth and rounded, and the ears are long and slender with a comparatively small number of rows or kernels. In temperate zones, flint corn often matures earlier, germinates better has more spring vigor, more tillers and fewer prop roots than dent strains.
Flour Corn
This is one of the oldest types of corn, tracing back to the ancient Aztecs and Incas. American and Indians used to ground the soft kernels for flour. Floury maize types have soft starch throughout, with practically no hard, vitreous endosperm and thus are opaque in kernel phenotype. Kernels tend to shrink uniformly upon drying, so usually have little or no denting. When dry, they are easy to grind, but may mold on the mature ear in wet areas.
Sweet Corn
In sweet corn, the sugary gene prevents or retards the normal conversion of sugar into starch during endosperm development, and the kernel accumulates a water-soluble polysaccharide called “phytoglycogen.” As a result, the dry, sugary kernels are wrinkled and glassy. The higher content of water-soluble polysaccharide adds a texture quality factor in addition to sweetness.
Popcorn
Popcorns are perhaps the most primitive of the surviving races of maize. This corn type is characterized by a very hard, corneous endosperm containing only a small portion of soft starch. Popcorns are essentially small-kernelled flint types. The kernels may be either pointed (rice-like) or round (pearl-like). Some of the more recently developed popcorns have thick pericarps (seed coats), while some primitive semi-popcorns, such as the Argentine popcorns, have thin pericarps.
Pod Corn
Pod corn (tunicate maize) is more of an ornamental type. The major gene involved (Tu) produces long glumes enclosing each kernel individually, which also occurs in many other grasses. The ear is also enclosed in husks, as with other types of corn. Homozygous pod corn usually is highly self-sterile. and the ordinary type of pod corn is heterozygous. Pod corn may be dent, sweet, waxy, pop, flint or floury in endosperm characteristics. It is merely a curiosity and is not grown commercially.
Waxy corn
Waxy corn name derives from the waxy appearance of the endosperm exposed in a cleanly cut cross-section. Common corn starch is approximately 73 percent amylopectin and 27 percent amylose, whereas waxy starch is composed entirely with amylopectin, which is the branched molecular form of starch. Ordinary corn starch stains blue with 2 percent potassium iodide solution, whereas waxy cornstarch stains a reddish brown. The waxy gene also expresses itself in the pollen with this staining reaction, which is an aid in breeding. Products made from waxy corn are used by the food industry as stabilizers and thickeners for puddings pie fillings, sauces, gravies, retorted foods, salad dressings, etc. Other waxy products are used as remoistening adhesives in the manufacture of gummed tape, in adhesives and in the paper industry. Waxy grain is also grown as a feed for dairy cattle and livestock.
High-amylose corn
Amylo-maize is the generic name for corn that has an amylose content higher than 50 percent. The endosperm mutant amylose extender (ae) was first observed by R. P. Bear in 1950, which increases the amylose content of the endosperm to about 60 percent in many dent backgrounds. Modifying factors alter the amylase contents as well as desirable agronomic characteristics of the grain. The amylose-extender gene expression is characterized by a tarnished, translucent, sometimes semifull kernel appearance. The starch from high-amylose corn is used in the textile industry, in gum candies (where its tendency to form a gel aids production), and as an adhesive in the manufacture of corrugated cardboard.
References
Brown, W.L. and Darrah, L.L. 1985. Origin, Adaptation, and Types of Corn. National Corn Handbook-10, pp.1-6.
Ritchie, S.W., Hanway, J.J. and Benson, G.O. 1993. How a Corn Plant Develops. Special Report No. 48. Iowa State University of Science and Technology Cooperative Extension Service Ames, Iowa.
Sleper, D.A. and Poehlman, J.M. (eds.). 2006. Breeding Corn (Maize); In Breeding Field Crops. Blackwell Publishing Professional 2121 State Avenue, Ames, Iowa, pp. 277-296.